【表現度発現の分子機構】
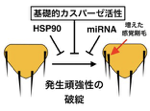
ある一つの遺伝子変異がもたらす表現型が異なる事例は数多く知られています。遺伝的な量的形質因子の関与や生育環境の違いによるエピゲノム制御がその要因として挙げられます。しかし、それらで説明がつかない事例も存在し、個体ごとの表現型の強弱といった違い(表現度:expressivity)を生み出す決定的な機構の研究はいまだ断片的です。遺伝子型が同じでも異なる表現型を生む機構で重要なものとしてプロテオスタシスがあり、HSP90シャペロンの関与が知られるが、我々は基質特異性の高いカスパーゼによる蛋白質プロセッシングが重要な仕組みとして機能する知見を得ています。我々は遺伝学的な解析に最適なショウジョウバエを用い、非細胞死性のカスパーゼ活性による特定の基質プロセッシング(Basal Caspase Processing: BCP)によって表現度を決めるメカニズムの理解からストレス応答や健康寿命を支える新たな仕組みの解明を行なっています。
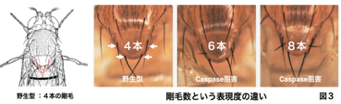
一般に発生の表現型は様々な遺伝的変異、あるいは環境刺激に対して頑強性をみせるが、少数の表現型に関しては同質遺伝子であっても表現度の違いを示します。ショウジョウバエカスパーゼ活性化因子(Adapf-1)変異体の解析から、胸部背盾板に左右2本ずつ生じる外感覚器である大剛毛の数が左右問わず増加しました(右図)。その分子メカニズムを調べた結果、感覚器前駆体細胞(SOP)が生まれるプロニューラルクラスターでBCPによってShaggy46/GSK3bのプロセッシングがおこって活性化し、モルフォゲンWntのシグナルを抑制することでSOP数を決めていました(Kanuka et al., EMBO J., 24, 3797, 2005)。このBCPは、自然免疫活性化やALS発症に関わるTBK1のショウジョウバエホモログIKKeによる内因性カスパーゼ阻害タンパク質DIAP1のリン酸化による分解促進によって調節されていました(Kuranaga et al., Cell 126,583, 2006)。大剛毛数は4本ですが野生型でも数%の割合で5~6本に増加する個体が現れます。この大剛毛数の増加を指標に関係する遺伝子のスクリーニングをした結果、遺伝子発現抑制性のヒストンメチル化であるH3K9me3を触媒するメチルトランスフェラーゼdSETDB1が得られました(Shinoda et al., GTC 21, 378, 2016)。カスパーゼ活性化変異体では大剛毛5~6本の個体の割合が40%に増加しますが、dSETDB1はカスパーゼ抑制とはパラレルに機能し、両方の抑制は大剛毛数増加個体の割合が90%に増加しました。重要な知見として内因性カスパーゼ阻害因子DIAP1ヘテロ変異体で、基礎的なカスパーゼ活性が高くなるとdSETDB1抑制による表現型が野生型に戻りました。これらの事実は表現度を生み出す仕組みとして、エピゲノム制御だけでなくそれとパラレルに働く新しい重要な仕組みとしてBCPがあることを示唆するものです。
ヒトや昆虫は左右相称動物(Bilateria)です。左右相性な生物においては個体の左右器官は発生時に同じ遺伝的背景と体内外環境を持つため、そのサイズや形態の非対称性の程度(これをFAと呼ぶ)は、個体内在性の生物学的ノイズに対する緩衝能力の低さを表すといえます。ヒトの左右の腕の長さをはかるとその正確さは驚異 的で0.2%しかぶれずFAは極めて低く抑えられています。ショウジョウバエ翅や脚も左右相称な組織で大きさに左右差はなくFAは低いことがわかりました。個体の健常性破綻時にはFAが増加していることも知られ(Am. J. Med. Genetics 66, 15, 1996; Cell Stress 12, 340, 2018)、疾患感受性を理解する観点からもFAの研究は重要です。カスパーゼ活性を発生期に阻害すると、予想に反して翅の細胞数が5-10%減少するとともに、左右の組織細胞数を一定に保つことができなくなり、翅のFAが増加することを見出しました(Shinoda et al., PNAS 116,20539,2019)。
これらの結果は発生時のBCPが感覚剛毛数、組織サイズ、FAという表現型の個体差すなわち表現度に寄与することを示しています。この現象に関わるBCPの分子機構の解明を行なっています。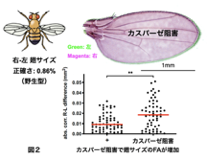
【文献】
Shinoda, N., Hanawa, N., Chihara, T., Koto, A., and *Miura, M.: Dronc-independent basal executioner caspase activity sustains Drosophila imaginal tissue growth.
Proc. Natl. Acad. Sci. USA., 116, 20539, 2019.
Shinoda, N., Obata, F., Zhang, L., and *Miura, M.: Drosophila SETDB1 and caspase cooperatively fine-tune cell fate determination of sensory organ precursor.
Genes to Cells 21, 378-386, 2016.
Kuranaga, E., Kanuka, H., Tonoki, A., Takemoto, K., Tomioka, T., Kobayashi, M., Hayashi, S., and *Miura, M.: Drosophila IKK-related kinase regulates nonapoptotic function of caspases via degradation of IAPs.
Cell 126, 583-596, 2006
Kanuka, H., Kuranaga, E., Takemoto, K., Hiratou, T., Okano, H., and *Miura, M.: Drosophila caspase transduces Shaggy/GSK-3b kinase activity in neural precursor development.
EMBO J. 24, 3793-3806, 2005
---------------------------------------------------------------------------------------------------------------------
This page is administered by Department of Genetics All Rights Reserved, Copyright © 2021 Department of Genetics
